alloreactivity - (Oct/15/2011 )
Dear all,
I am having troubles with understanding the principles behind alloreactivity or tissue-rejection.
How I understand it:
When an organ/tissue is transplanted you will have foreign MHC molecules of this foreign tissue that will be recognised by acceptor T-cells.
This happens because the foreign MHC molecules are "similar" to the own MHC molecules.
This means :
That the T cells recognise the foreign MHC with a strange peptide in it? (endogenous/exogenous peptide, where does does peptide come from?).
(this means you have a "regular MHC-Tcell reaction" , meaning T cells recognises strange MHC molecule as self MHC molecule + recognises strange peptide in it)
Or and here is where I get stuck: Do the own T cells recognise these foreign MHC molecules as an antigen on itself (meaning: foreign MHC molecule is seen as a self MHC + strange peptide, thus a piece of the foreign MHC will be seen as a strange peptide).
(I do know about the indirect pathway which means that the APC can present a piece of strange MHC that has been broken down, so this is not a problem for me)
Another thing where I get stuck is why the donor organ gerts destroyed.
I can understand the whole concept of recognition of foreign MHC etc... but why is the donor organ being destroyed by this? I mean: in the entire body you have MCH - Tcell reactions and this does not end up by having tissue destroyed.. So how come the donor tissue gets destroyed?
(simply put: own body parts have MHC molecules too, but how come this does not result in tissue destruction?)
any help appreciated
I will try to give my view of things. There are two types of recognition (see attached image) and in case of host versus graft rejection they are both because of activation of the host's T cells. They can be either activated against the donor's MHC molecules in the graft or peptides that are produced from the grafted tissue that bind to the host's MHC molecules and activate the immune response.
To answer your last question. Both types of these reactions happen only in the grafted tissue, because these molecules (either allogeneic MHC molecules or graft-derived peptides) are found only in the grafted tissue.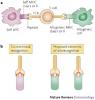
BioMiha on Sun Oct 16 09:03:31 2011 said:
I will try to give my view of things. There are two types of recognition (see attached image) and in case of host versus graft rejection they are both because of activation of the host's T cells. They can be either activated against the donor's MHC molecules in the graft or peptides that are produced from the grafted tissue that bind to the host's MHC molecules and activate the immune response.
To answer your last question. Both types of these reactions happen only in the grafted tissue, because these molecules (either allogeneic MHC molecules or graft-derived peptides) are found only in the grafted tissue.
Thanks.
But I understand this.
What I wonder about is: how come the "attack" of the immune system against the graft is more severe then the regular attacks to the acceptor his own tissue.
I understand that the graft only has allogeneic MHC or graft derived peptides, but other parts of the acceptor body has own MHC molecules..
Or are there (for some reason) more MHC molecules in the graft then in regular tissue?
PS. when you say: " donor's MHC molecules" , do you mean that the acceptor his Tcells respond to the MHC moluces itself? (meaning there does not need to be a peptide in it? ==> the MHC molecules plays both the role of "own MHC molecule" and "strabge peptide in it")
Why would the acceptor attack his own tissue? That's the whole point of tolerance to self... I mean sometimes it happens but in that case we call it autoimmune disease. The body has a multitude of mechanisms to control autoreactivity.
And no, the MHC molecules are not stable if there is no peptide bound. The image I attached to my previous post is really descriptive about where the TCR binds.
BioMiha on Sun Oct 16 10:20:38 2011 said:
Why would the acceptor attack his own tissue? That's the whole point of tolerance to self... I mean sometimes it happens but in that case we call it autoimmune disease. The body has a multitude of mechanisms to control autoreactivity.
And no, the MHC molecules are not stable if there is no peptide bound. The image I attached to my previous post is really descriptive about where the TCR binds.
What I ment is: on the acceptor his own tissue are also MHC molecules with strange peptides... => so why is this tissue not destroyed... (there must be a reason why there is a difference between the "attack" on the strange tissue and the own tissue)
How I see it:
Own MHC + strange peptide = attack of T cells
Or: strange MHC (donor) is seen as own MHC + strange peptide in it (as you mention that there has to be a peptide in it) = attack of T cells.
Now: how come that in the first case your own tissue is not destroyed and in the second case the donor tissue is destroyed.
About the picture: if you watch at the proposed extremes, it does look like that the T cell receptor can react on a strange pepite alone (and not make contact with the MHC) or that it can react with the strange MCH (and does not need to bind with the peptide) ==> this makes me wonder: why do our own T cells bind with strange MHC whilst not binding the Peptide in it or why does it bind the strange peptide alone and not make contact with the MHC, because we all know that T cells normalle need to make contact with the MHC molecule and the peptide.
The own tissue of the recipient has self MHC molecules + self peptides (not strange or foreign as you say) that is why it does not get attacked. If for example there are foreign peptides in that tissue, e.g. from a bacterium or virus, those cells get attacked, because that is how the immune system works.
Concerning the image: the TCR can bind either the foreing MHC with self peptide or self MHC with foreign peptide or foreign MHC with foreign peptide (which is probably a rare event). Where it binds is not critical for immune system activation.
BioMiha on Sun Oct 16 13:32:24 2011 said:
The own tissue of the recipient has self MHC molecules + self peptides (not strange or foreign as you say) that is why it does not get attacked. If for example there are foreign peptides in that tissue, e.g. from a bacterium or virus, those cells get attacked, because that is how the immune system works.
Concerning the image: the TCR can bind either the foreing MHC with self peptide or self MHC with foreign peptide or foreign MHC with foreign peptide (which is probably a rare event). Where it binds is not critical for immune system activation.
I understand what you mean however the question still stays the same: How come when the self tissue shows MHC with strange peptides (from bacteria for example) is not destroyed by the action of the Tcells?
(only some cells get destroyed, but the tissue on itself survives.. there must be a reason why the attack against "donor" tissue is stronger then against own tissue. And this is what I do not know.)
About the image: you say that its rare that it can bind a foreign MHC with foreign peptide.. but really? is this true? I dont think so, because this is just the problem with tissue rejection: binding of foreign MHC with foreign peptide...
(PS.: I have always been told that MHC class 1 with self peptides in it do NOT leave the intern area of the cell: it does not move to the golgi complex and thus does not go to the exterior of the cell.. this means that on our own cells there are normally no MHC 1 with self peptides ==> what I want to say with this is that the own tissue does not really have MHC molecules with self peptides on the outside, so off course it does not get attacked ... but if it does: then why does the tissue surive)
Honestly, I think you are preoccupied by your preconceptions and are unwilling to look at things from a different perspective.
I will give my best to answer your questions, however I suggest you take a look at Abbas: Cellular and Molecular Immunology.
When cells of a certain tissue are infected (by bacteria, viruses, etc...) CD8 T cells kill the infected cells because these cells present foreign peptides bound to self MHC molecules. These cells die by apoptosis and are replaced by new cells. If on the other hand, a large number of cells is infected, the immune activation can indeed destroy a lot of tissue, which is replaced by scar tissue (e.g. in case of tuberculosis). In the case of graft rejection EVERY cell of the graft is foreign to the recipients body and is therefore targeted for destruction that is why the whole graft is rejected. There is no fundamental difference in the way the immune system is activated. If for example every cell of your own tissue were infected with a virus, it too would be attacked in the same way. However, there is a big difference in the magnitude of the reaction, because in most infections the body has a great capacity to limit the spread of the infection and therefore most cells don't get infected and the tissue survives.
For your second question. MHC molecules bind peptides in a random manner. That means that they also bind self peptides and present them to T cells. Potentially self reactive T cells are removed in the process of T cell development in the thymus so the the remaining T cells do not bind to self MHC + self peptide complexes with a high enough affinity to get activated (or in the majority of cases if they do bind it is in the absence of co-stimmulatory molecules, so these cells become anergic). I don't know who told you that MHC class I molecules do not present self peptides but this is just wrong.
For your second question. MHC molecules bind peptides in a random manner. That means that they also bind self peptides and present them to T cells. Potentially self reactive T cells are removed in the process of T cell development in the thymus so the the remaining T cells do not bind to self MHC + self peptide complexes with a high enough affinity to get activated (or in the majority of cases if they do bind it is in the absence of co-stimmulatory molecules, so these cells become anergic). I don't know who told you that MHC class I molecules do not present self peptides but this is just wrong.
Ok, I get it now.
This is why I couldnt understand it: someone (a teacher) did tell me that there only would be MHC1 molecules on cells when they had foreign peptides on them! So I couldnt figure out why the donor tissue would be destroyed.
But ofcourse, if they present self peptides (peptides from the donor) then the acceptor would indeed see those peptides as foreign and attack and since there are so many it would indeed cause a massive attack.
Just one more question:
Do you know why the (or how) the body is able to limit the spread of the infection in the own tissue and not in the donor tissue?
Thanks a lot allready, I think I understand it now
lucilius on Mon Oct 17 07:02:27 2011 said:
Do you know why the (or how) the body is able to limit the spread of the infection in the own tissue and not in the donor tissue?
Thanks a lot allready, I think I understand it now
In case of the graft there is no infection - there is no spreading. The cells of the graft synthesize the molecules by themselves and present them to the recipients immune system. And because all of the cells of the graft are foreign they are all targets for the CD8 cells.